Нейронные механизмы и воздействие на них. часть 1
Содержание:
Взаимодействие с нейромедиаторами
Нейроны разного местонахождения общаются между собой с помощью электрических импульсов химической природы. Так, что же лежит в основе их образования? Существуют так называемые нейромедиаторы (нейротрансмиттеры) – сложные химические соединения. На поверхности аксона располагается нервный синапс – контактная поверхность. С одной стороны находится пресинаптическая щель, а с другой – постсинаптическая. Между ними находится щель – это и есть синапс. На пресинаптической части рецептора располагаются мешочки (везикулы), содержащие определенное количество нейромедиаторов (квант).
Когда импульс подходит к первой части синапса, инициируется сложный биохимический каскадный механизм, в результате которого мешочки с медиаторами вскрываются, и кванты веществ-посредников плавно вытекают в щель. На этом этапе импульс исчезает, и появляется вновь только тогда, когда нейромедиаторы достигают постсинаптической щели. Тогда снова активируются биохимические процессы с открытиями ворот для медиаторов и те, действуя на мельчайшие рецепторы, преобразуются в электрический импульс, идущий далее в глубины нервных волокон.
Между тем выделяют разные группы этих самых нейромедиаторов, а именно:
- Тормозные нейромедиаторы – группа веществ, осуществляющие тормозное действие на возбуждение. К ним относят:
- гамма-аминомасляную кислоту (ГАМК);
- глицин.
- Возбуждающие медиаторы:
- ацетилхолин;
- дофамин;
- серотонин;
- норадреналин;
- адреналин.
Пресинаптическая часть
Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором, элементы цитоскелета и митохондрии. В пресинаптическую мембрану встроены потенциалозависимые Ca2+?каналы. При поступлении ПД к терминальному расширению мембрана деполяризуется, Ca2+?каналы открываются, ионы Ca2+ входят в терминаль, запуская в активных зонах процесс слияния мембраны синаптического пузырька и пресинаптической мембраны, т.е. секрецию (экзоцитоз) нейромедиатора (рис. 6–6, позиции 2–4).
Роль Са2+. Слияние синаптических пузырьков с пресинаптической мембраной происходит при увеличении концентрации Са2+ в цитозоле нервной терминали. Белок синаптического пузырька синаптотагмин связывается с Са2+ и тем самым принимает участие в регуляции экзоцитоза (в том числе путём реорганизации примембранного цитоскелета).
Синаптические пузырьки. Молекулы нейромедиатора накапливаются в нервной терминали, находясь внутри синаптических пузырьков вместе с АТФ и некоторыми катионами. В каждом пузырьке находится несколько тысяч молекул нейромедиатора, что составляет квант нейромедиатора.
Синтез нейромедиатора. Ферменты, необходимые для образования нейромедиаторов, синтезируются в перикарионе и транспортируются к синаптической терминали по аксонам (рис. 6–4).
Типы пузырьков — мелкие (диаметр порядка 50 нм) и крупные (диаметр 100–200 нм). Мелкие синаптические пузырьки содержат «классические» медиаторы (см. ниже). Крупные везикулы содержат нейропептиды.
Секреция. Когда ПД достигает нервной терминали, синаптические пузырьки сливаются с пресинаптической мембраной, что приводит к выделению квантов нейромедиатора в синаптическую щель. Незначительное количество квантов нейромедиатора постоянно (спонтанно) секретируется в синаптическую щель.
Узнавание. Предшествующий слиянию синаптических пузырьков и плазмолеммы процесс узнавания синаптическим пузырьком пресинаптической мембраны происходит при взаимодействии мембранных белков (синаптобревин, SNAP-25, синтаксин и другие).
Влияние токсинов. Синтаксин, SNAP-25 и синаптобревин — мишени ботулинического токсина, необратимо подавляющего слияние синаптических пузырьков с пресинаптической мембраной. Мишень столбнячного токсина — синаптобревин.
Активные зоны (рис. 6–3). Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания — активных зонах — участках утолщения пресинаптической мембраны. Активная зона состоит из «плотной полоски» на пресинаптической мембране и сгруппированных около неё синаптических пузырьков, потенциалозависимых кальциевых каналов, специальных белков экзоцитоза и элементов цитоскелета. Количество активных зон в нервно-мышечном синапсе достигает 30–40, в межнейронных синапсах — около десятка. Активные зоны расположены против скоплений рецепторов в постсинаптической мембране, что уменьшает задержку в передаче сигнала, связанную с диффузией нейромедиатора в синаптической щели.
Рис. 6-3. Активные зоны нервно-мышечного синапса расположены напротив постсинаптических складок — участков скоплений холинорецепторов. Пресинаптическая мембрана слева расщеплена на два листка.
Жизненный цикл синаптических пузырьков (рис. 6–4). Синаптические везикулы образуются в теле нейрона в эндоплазматическом ретикулуме и комплексе Гольджи (1) и с аксонным транспортом поступают в нервные окончания (2).
Рис. 6-4. Образование, транспорт и экзоцитоз синаптических пузырьков.
В нервном окончании мелкие синаптические пузырьки посредством активного транспорта заполняются медиатором (3) и передвигаются к пресинаптической мембране (4). Освобождение медиатора (5) может осуществляться посредством экзоцитоза с полным («классический» механизм) либо неполным (механизм «kiss and run») слиянием. Первый вид экзоцитоза сопровождается встраиванием мембраны везикулы в пресинаптическую, опорожнением пузырька, а затем посредством эндоцитоза образуются покрытые клатрином везикулы (6), которые затем проходят стадию эндосомы (7) и снова заполняются медиатором (3). Второй вид экзоцитоза характеризуется образованием временной поры, соединяющей полость пузырька с синаптической щелью. После выделения медиатора везикула не встраивается в пресинаптическую мембрану, а отпочковывается от неё (8) и повторно заполняется медиатором (3). Крупные синаптические везикулы заполняются медиатором в теле клетки (9), их экзоцитоз происходит в других участках пресинаптической мембраны, а эндоцитоз опорожнённых пузырьков отсутствует (10).
-
Назад
-
Вперёд
Ваша роль в мозговом общении
Если аксоны ответственны за то, что нервные импульсы проходят через две точки нервной системы, дендриты отвечают за захват химических веществ, которые выходят из кончика аксонов и сделать эти химические сигналы преобразовывать или нет в электрические импульсы Хотя этот процесс также может быть инициирован в теле нейрона.
То есть именно в дендритах и в нейронном теле, где рождаются электрические сигналы (также называемые потенциалами действия), которые проходят через нейроны и заканчиваются на кончике аксонов, заставляя эту часть нейрона выделять химические вещества. Когда нужное количество нейротрансмиттеров достигает дендритов, происходит деполяризация , который является процессом, который генерирует нервные импульсы.
Дендриты Они очень чувствительны к малейшим изменениям типа и количества нейротрансмиттеров, которые они собирают и это означает, что в зависимости от химических веществ, которые они обнаруживают, они инициируют ту или иную комбинацию электрических импульсов или что электрический сигнал не генерируется напрямую, если выполняются условия.
Это означает, что необязательно, чтобы дендриты не улавливали какой-либо нейротрансмиттер, чтобы они не производили электрический импульс ; Это также может произойти, если они улавливают определенное количество определенного типа химического вещества.Вот почему некоторые психотропные препараты воздействуют на дендриты нейронов, заставляя их не генерировать электрические сигналы, как если бы не было эффекта этого активного начала.
Короче говоря, молекулярные следы, оставленные живым опытом в дендритах и терминалах нейронов, являются основой функционирования нервной системы и ее способности динамически изменять свою активность. В то же время они являются фундаментальной частью процесса управления памятью, которые представляют собой напечатанные узоры в тех молекулярных следах, с которыми работает нервная клетка.
Рост и развитие аксона[править]
Нейрон
Рост аксонов происходит через их окружающую среду, в виде конуса роста, который находится в наконечнике аксона. Конус роста имеет широкий лист как расширение, названное lamellipodia, которое содержат выпячивания, названные filopodia. Filopodia — механизм, представляющий процесс придержки поверхностей. Он анализирует ближайшую окружающую среду. Актин играет главную роль в подвижности этой системы. Окружающие среды с высокими уровнями молекул прилипания ячейки или «КУЛАКА» создают идеальную окружающую среду для аксонального роста. Это, кажется, обеспечивает «липкую» поверхность для аксонов, для раста вперед. Примеры КУЛАКА, определенного для нервных систем включают: N-КУЛАК, neuroglial КУЛАК или NgCAM, ПОМЕТЬТЕ 1, МЭГ, и DCC, все из которых — часть суперсемьи иммуноглобулина. Другой набор молекул звонковый, внеклеточные матричные молекулы прилипания также обеспечивают липкое основание для аксонов, чтобы расти вперед. Примеры этих молекул включают laminin, fibronectin, tenascin, и perlecan. Некоторые из них — поверхность, привязанная к ячейкам и таким образом действуют, как короткие аттрактанты диапазона или repellents. Другие — difusible лиганды и таким образом могут долго сохранять эффекты диапазона.
Ячейки звонковые, ячейки указательного столба помогают в руководстве ростом аксона нейронала. Эти ячейки — типично другой, иногда незрелый, нейроны.
Тело клетки
Тело
нервной клетки состоит из протоплазмы(цитоплазмыиядра),
снаружи ограничена мембраной из двойного
слоялипидов(билипидный слой). Липиды состоят изгидрофильныхголовок и гидрофобных хвостов, расположеныгидрофобнымихвостами друг к другу, образуягидрофобныйслой, который пропускает только
жирорастворимые вещества (напр. кислород
и углекислый газ). На мембране находятся
белки: на поверхности (в форме глобул),
на которых можно наблюдать наросты
полисахаридов (гликокаликс), благодаря
которым клетка воспринимает внешнее
раздражение, и интегральные белки,
пронизывающие мембрану насквозь, в
которых находятся ионные каналы.
Несмотря на это ограничение, персептроны с порогом обладают интересным свойством, которое мы рассмотрим в следующем разделе: существует алгоритм, позволяющий персептрону адаптировать свои веса к набору примеров, чтобы получить для этого вместе ожидаемая классификация. Таким образом, если набор примеров достаточно велик, можно получить персептрон, который даст подходящие результаты для ненаблюдаемых примеров.
Существует два алгоритма, в основном, для «обучения» монослойной нейронной сети. Первый — это простой метод и называется градиентным спусками. Эти два метода состоят в сравнении результата, который ожидался для примеров, а затем в минимизации ошибки, сделанной на примерах, но, конечно же, существует разница между двумя методами, которые будут объяснены ниже.
Нейрон
состоит из тела диаметром от 3 до 130 мкм,
содержащего ядро (с большим количеством
ядерных пор) и органеллы (в том числе
сильно развитый шероховатый ЭПРс активнымирибосомами,аппарат
Гольджи), а также из отростков. Выделяют
два вида отростков: дендриты и аксон.
Нейрон имеет развитый и сложный
цитоскелет, проникающий в его отростки.
Цитоскелет поддерживает форму клетки,
его нити служат «рельсами» для транспорта
органелл и упакованных в мембранные
пузырьки веществ (например, нейромедиаторов).
Цитоскелет нейрона состоит из фибрилл
разного диаметра:Микротрубочки(Д = 20-30 нм) — состоят из белкатубулинаи тянутся от нейрона по аксону, вплоть
до нервных окончаний. Нейрофиламенты
(Д = 10 нм) — вместе с микротрубочками
обеспечивают внутриклеточный транспорт
веществ. Микрофиламенты (Д = 5 нм) —
состоят из белков актина и миозина,
особенно выражены в растущих нервных
отростках и в нейроглии. В теле нейрона
выявляется развитый синтетический
аппарат, гранулярная ЭПС нейрона
окрашиваетсябазофильнои известна под названием «тигроид».
Тигроид проникает в начальные отделы
дендритов, но располагается на заметном
расстоянии от начала аксона, что служит
гистологическим признаком аксона.
Нейроны различаются по форме, числу
отростков и функциям. В зависимости от
функции выделяют чувствительные,
эффекторные(двигательные, секреторные)
и вставочные. Чувствительные нейроны
воспринимают раздражения, преобразуют
их в нервные импульсы и передают в мозг.
Эффекторные (от лат. эффектус —
действие) — вырабатывают и посылают
команды к рабочим органам. Вставочные —
осуществляют связь между чувствительными
и двигательными нейронами, участвуют
в обработке информации и выработке
команд.
Обучение градиентным спуском
Мы будем, по каждому из методов, изучать коррекцию весов только для одного из нейронов, достаточно будет применить метод по вашему выбору к каждому из нейронов монослоевой сети. Таким образом, мы видим, что ошибка равна нулю, если нейронная сеть не является неправильной в любом из примеров, то есть, если она правильно рассчитает правильный вывод для каждого из примеров. мы начинаем с взвешенных выстрелов случайным образом. Однако, поскольку результат очень важен, демонстрация доступна в приложении к этой статье.
Различается
антероградный (от тела) и ретроградный
(к телу) аксонный транспорт.
Сложная обёртка
По сути миелин — это клеточная мембрана глиальных клеток, многократно обмотанная вокруг аксона. Сама мембрана на 70–75% состоит из липидов и на 25–30% — из белков. В периферической нервной системе донором мембран становятся , а в центральной — . Эти клетки бережно обматывают своими мембранами ценные каналы связи, чтобы обеспечить надежное взаимодействие нервной системы и периферических органов. Миелин покрывает нервное волокно не целиком: существуют промежутки между наслоениями миелина, называемые перехватами Ранвье (рис. 1). Есть прямая зависимость между расстоянием от одного промежутка до другого и скоростью распространения нервного импульса по волокну: чем больше расстояние между перехватами Ранвье, тем выше скорость передачи сигнала в нерве .
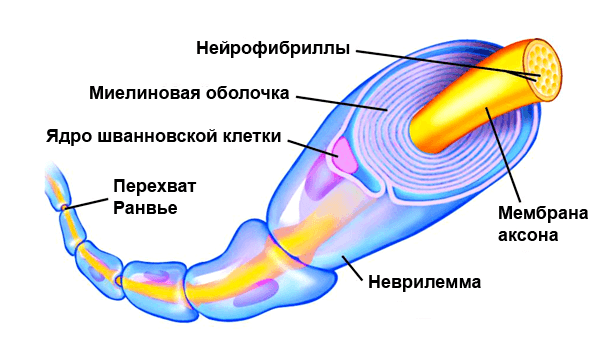
Рисунок 1. Нервное волокно, обернутое миелином. Видны ядра шванновских клеток (nucleus of Schwann cell) и перехваты Ранвье (nodes of Ranvier) — участки аксона, которые не покрыты миелиновой оболочкой.
Если говорить о белках, входящих в состав миелина, то надо уточнить, что это не только простые белки. В миелине встречаются гликопротеины — белки, к которым присоединены короткие углеводные последовательности
Важной составляющей миелина является главный структурный белок миелина (myelin basic protein, MBP), впервые выделенный около 50 лет назад. MBP — это трансмембранный белок, который может многократно «прошивать» липидный слой клетки
Его различные изоформы (рис. 2) кодируются геном под названием Golli (gene in the oligodendrocyte lineage). Структурной основой миелина служит изоформа массой 18,5 килодальтон .
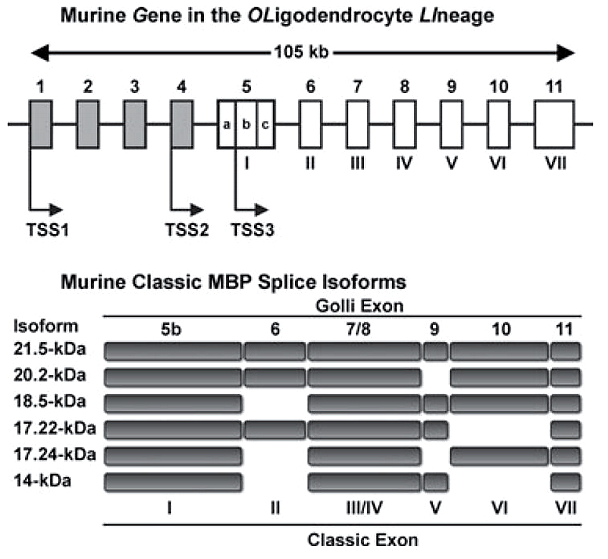
Рисунок 2. Различные изоформы основного белка миелина (MBP) создаются на основе одного и того же гена. Например, для синтеза изоформы массой 18,5 кДа используются все экзоны, кроме экзона II.