Синапс
Содержание:
История открытия
- В 1897 году Шеррингтон сформулировал представление о синапсах.
- За исследования нервной системы, в том числе синаптической передачи, в 1906 году Нобелевскую премию получили Гольджи и Рамон-и-Кахаль.
- В 1921 австрийский учёный О. Лёви (О. Loewi) установил химическую природу передачи возбуждения через синапсы и роль в ней ацетилхолина. Получил Нобелевскую премию в 1936 г. совместно с Г. Дейлом (Н. Dale).
- В 1933 советский учёный А. В. Кибяков установил роль адреналина в синаптической передаче.
- 1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) получили Нобелевскую премию за открытие роли норадреналина в синаптической передаче.
Пресинаптическая часть
Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором, элементы цитоскелета и митохондрии. В пресинаптическую мембрану встроены потенциалозависимые Ca2+?каналы. При поступлении ПД к терминальному расширению мембрана деполяризуется, Ca2+?каналы открываются, ионы Ca2+ входят в терминаль, запуская в активных зонах процесс слияния мембраны синаптического пузырька и пресинаптической мембраны, т.е. секрецию (экзоцитоз) нейромедиатора (рис. 6–6, позиции 2–4).
Роль Са2+. Слияние синаптических пузырьков с пресинаптической мембраной происходит при увеличении концентрации Са2+ в цитозоле нервной терминали. Белок синаптического пузырька синаптотагмин связывается с Са2+ и тем самым принимает участие в регуляции экзоцитоза (в том числе путём реорганизации примембранного цитоскелета).
Синаптические пузырьки. Молекулы нейромедиатора накапливаются в нервной терминали, находясь внутри синаптических пузырьков вместе с АТФ и некоторыми катионами. В каждом пузырьке находится несколько тысяч молекул нейромедиатора, что составляет квант нейромедиатора.
Синтез нейромедиатора. Ферменты, необходимые для образования нейромедиаторов, синтезируются в перикарионе и транспортируются к синаптической терминали по аксонам (рис. 6–4).
Типы пузырьков — мелкие (диаметр порядка 50 нм) и крупные (диаметр 100–200 нм). Мелкие синаптические пузырьки содержат «классические» медиаторы (см. ниже). Крупные везикулы содержат нейропептиды.
Секреция. Когда ПД достигает нервной терминали, синаптические пузырьки сливаются с пресинаптической мембраной, что приводит к выделению квантов нейромедиатора в синаптическую щель. Незначительное количество квантов нейромедиатора постоянно (спонтанно) секретируется в синаптическую щель.
Узнавание. Предшествующий слиянию синаптических пузырьков и плазмолеммы процесс узнавания синаптическим пузырьком пресинаптической мембраны происходит при взаимодействии мембранных белков (синаптобревин, SNAP-25, синтаксин и другие).
Влияние токсинов. Синтаксин, SNAP-25 и синаптобревин — мишени ботулинического токсина, необратимо подавляющего слияние синаптических пузырьков с пресинаптической мембраной. Мишень столбнячного токсина — синаптобревин.
Активные зоны (рис. 6–3). Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания — активных зонах — участках утолщения пресинаптической мембраны. Активная зона состоит из «плотной полоски» на пресинаптической мембране и сгруппированных около неё синаптических пузырьков, потенциалозависимых кальциевых каналов, специальных белков экзоцитоза и элементов цитоскелета. Количество активных зон в нервно-мышечном синапсе достигает 30–40, в межнейронных синапсах — около десятка. Активные зоны расположены против скоплений рецепторов в постсинаптической мембране, что уменьшает задержку в передаче сигнала, связанную с диффузией нейромедиатора в синаптической щели.
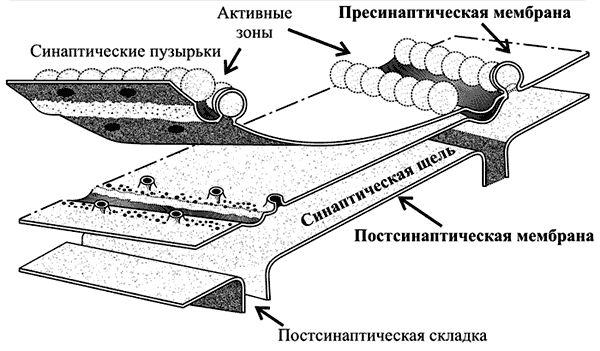
Рис. 6-3. Активные зоны нервно-мышечного синапса расположены напротив постсинаптических складок — участков скоплений холинорецепторов. Пресинаптическая мембрана слева расщеплена на два листка.
Жизненный цикл синаптических пузырьков (рис. 6–4). Синаптические везикулы образуются в теле нейрона в эндоплазматическом ретикулуме и комплексе Гольджи (1) и с аксонным транспортом поступают в нервные окончания (2).
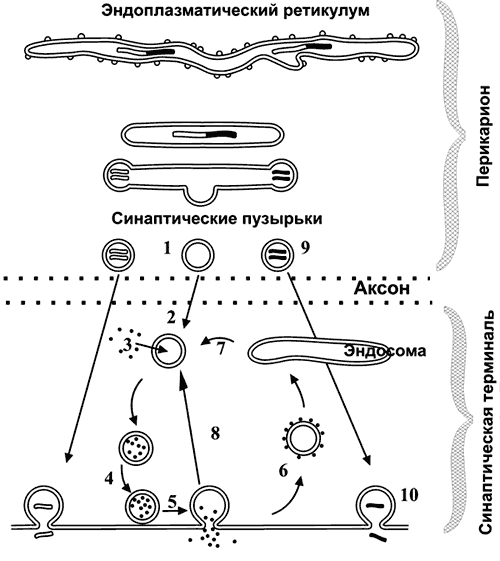
Рис. 6-4. Образование, транспорт и экзоцитоз синаптических пузырьков.
В нервном окончании мелкие синаптические пузырьки посредством активного транспорта заполняются медиатором (3) и передвигаются к пресинаптической мембране (4). Освобождение медиатора (5) может осуществляться посредством экзоцитоза с полным («классический» механизм) либо неполным (механизм «kiss and run») слиянием. Первый вид экзоцитоза сопровождается встраиванием мембраны везикулы в пресинаптическую, опорожнением пузырька, а затем посредством эндоцитоза образуются покрытые клатрином везикулы (6), которые затем проходят стадию эндосомы (7) и снова заполняются медиатором (3). Второй вид экзоцитоза характеризуется образованием временной поры, соединяющей полость пузырька с синаптической щелью. После выделения медиатора везикула не встраивается в пресинаптическую мембрану, а отпочковывается от неё (8) и повторно заполняется медиатором (3). Крупные синаптические везикулы заполняются медиатором в теле клетки (9), их экзоцитоз происходит в других участках пресинаптической мембраны, а эндоцитоз опорожнённых пузырьков отсутствует (10).
История изучения
Впервые гипотеза о существовании электрических синапсов была выдвинута в начале XX века Камилло Гольджи и Йозефом Герлахом. Однако после открытия химических синапсов существование электрических синапсов считалось недостоверным, и до середины XX века главенствовало мнение, что передача потенциала действия между нейронами осуществляется исключительно посредством химических синапсов. Тем не менее, в 1959 году Дэвид Поттер и Эдвин Фершпан убедительно доказали существование электрических синапсов на примере гигантского аксона и аксоном моторного нейрона в брюшной нервной цепочке рака.
Распространение
Электрические синапсы встречаются по всей центральной нервной системе. Они были детально изучены в неокортексе, гиппокампе, , голубом пятне, , , обонятельных луковицах, сетчатке и спинном мозге позвоночных. Электрические синапсы были обнаружены также в полосатых телах, мозжечке и супрахиазматическом ядре. Как правило, электрические синапсы опосредуют очень быстрые поведенческие ответы, как, например, выпускание чернил морским зайцем Aplysia при опасности. Электрические синапсы были обнаружены и вне нервной системы. Электрические синапсы связывают , кардиомиоциты, гладкомышечные клетки, клетки печени, глиальные и эпителиальные клетки.
Электрические синапсы наиболее характерны для низкоорганизованных животных. В ходе эволюции доля электрических синапсов уменьшалась, и в ЦНС млекопитающих (в том числе человека) на долю электрических синапсов приходится около 1 % связей между нейронами.
Классификации синапсов
Основные элементы электрического синапса (эфапса): а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона
По механизму передачи нервного импульса
- химический — это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
- электрический (эфапс) — место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований — коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм). Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
- смешанные синапсы — пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
По местоположению и принадлежности структурам
- периферические
- нервно-мышечные
- нейросекреторные (аксо-вазальные)
- рецепторно-нейрональные
- центральные
-
аксо-дендритические — с дендритами
аксо-шипиковые — с дендритными шипиками, выростами на дендритах;
, в том числе
- аксо-соматические — с телами нейронов;
- аксо-аксональные — между аксонами;
- дендро-дендритические — между дендритами;
-
аксо-дендритические — с дендритами
Различные варианты расположения химических синапсов
По нейромедиатору
- аминергические, содержащие биогенные амины (например, серотонин, дофамин
в том числе адренергические, содержащие адреналин или норадреналин;
);
- холинергические, содержащие ацетилхолин;
- пуринергические, содержащие пурины;
- пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия синапса
- возбуждающие
- тормозные.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов:
1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала;
2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Строение синапса и его классификации
Все синапсы состоят из трёх основных элементов:
- Пресинаптическая мембрана – образована из окончаний конечных ветвлений аксона или дендрита (в дендродендритном синапсе). Покрывает расширенное нервное окончание – нейросекреторный аппарат. Синтез медиатора происходит благодаря пузырькам и митохондриям, что находятся в пресинаптической части. Медиаторы содержатся в гранулах, или пузырьках.
- Синаптическая щель – пространство между мембранами пресс- и постсинаптической мембранами, что заполнены жидкостью, которая похожа по структуре и составу на плазму крови. Щель разделяет аксон одной клетки и дендрит другой клетки. Именно через эту прорезь и получает последующая клетка импульс от предыдущей клетки (от 20 до 30 нанометров).
- Постсинаптическая мембрана – представляет собой утолщённую часть мембраны клетки, которая и контактирует с пресинаптической мембраной, т.е. получает импульс. Она имеет специальные ионные каналы, что создают потенциал действия (возбуждение, что перемещается по мембране ЖИВОЙ клетки в процессе прохождения нервного сигнала). На ней имеются специфические белковые организации, что исполняют роль рецепторов, которые воспринимают действие медиаторов.
Виды синапсов и их классификация
Синапсы можно систематизировать:
Классификация по механизму:
- Электрические;
- Химические;
- Классификация по росту в онтогенезе:
- Стабильные – синапсы дуг врожденных рефлексов;
- Динамичные – в процессе формирования, т.е. преобретенных рефлексов.
Классификация по медиатору, с помощью которого передаётся импульс:
- Холинергические – медиатор с АЦХ (ацитилхолин);
- Серотонинергические – медиатор с биогенным амином, т.е. с серотонином.
Синапсы можно систематизировать:
Химические синапсы
В химических синапсах импульс передаётся посредством медиатора, т.е. проводника. Синапсы ЦНС имеют различное строение, классификацию и функциональные свойства.
Медиатор – биологически активные вещества, что секретируются на концах отростков нервных волокон и позволяют пройти нервному импульсу в синапсах.
Свойства:
- Удвоение (репликация) возникает в нейроне,
- Скопление в конце клетки,
- Производство при появление иона Са2 в пресинаптическом конце.
- Биогенные амины (Гистамин),
- Аминокислоты (Таурин),
- Пурины (АТФ, аденозин),
- Нейропептиды (Нейротензин),
- Газы (H2S, NO, CO).
Протекает в четыре этапа:
- Изготовление медиатора.
- Возникает в пресинаптическом конце, или в теле нейрона. Это происходит только при непосредственном участии катализаторов – ферментов.
- Когда синтез проходит в теле нейрона, нужны пузырьки оболочки – мембраны – которые создаёт Комплекс Гольджи – везикулы – они по микротрубочкам переходят в пресинаптическое окончание. У любого из нейронов есть 1 главный медиатор и несколько вспомогательных.
- Выход медиатора в синаптическое пространство (щель).
Идёт в результате воздействия появившегося возбуждения. Это приводит к усилению проницаемости мембраны для ионов Ca2 , которые выходят в цитоплазму постсинаптического окончания. Именно ионы 2 кальция и обеспечивают изменение белков мембран везикул (пузырьков) из-за чего пузырьки и перемещаются к пресинаптической мембране. Секреция медиатора происходит за 1-5 мс. Содержимое одного кванта медиатора составляет около 104 молекул медиатора.
Медиатор согласуется с рецептором постсинаптической мембраны, что вызывает его возбуждение. С помощью диффузии молекулы медиатора проходят сквозь щель, после чего достигают постсинаптической мембраны, которая способна «распознать» свой медиатор с помощью специальных рецепторов.
Электрические синапсы.
Главное различие электрических синапсов от химических – очень малая синаптическая щель и небольшая электрическая устойчивость между стенками. Эти синапсы характеры для схожих – однотипных – клеток. Импульс запросто проходит через мембраны. Электрические синапсы бывают с одно- и двусторонней трансмиссией возбуждения.
Одинаковые свойства:
- Быстрый эффект,
- Практически невозможна суммация последовательных сигналов – слабость следовых эффектов,
- Высокая надежность передачи возбуждения.
- Ток из пресипнаптического потенциала действия раздражает постсинаптическую мембрану, где возникает потенциал действия. Характерно для сердечной мышцы.
- Наряду с электрическими синапсами существуют синапсы тормозного действия – влияние пресинаптической нервной клетки на постсинаптическую. Это сопровождается устранение процесса возбуждения. Тормозных синапсов в ЦНС больше, чем возбуждающих.
История открытия
- В 1897 году Шеррингтон сформулировал представление о синапсах.
- За исследования нервной системы, в том числе синаптической передачи, в 1906 году Нобелевскую премию получили Гольджи и Рамон-и-Кахаль.
- В 1921 австрийский учёный О. Лёви (О. Loewi) установил химическую природу передачи возбуждения через синапсы и роль в ней ацетилхолина. Получил Нобелевскую премию в 1936 г. совместно с Г. Дейлом (Н. Dale).
- В 1933 советский учёный А. В. Кибяков установил роль адреналина в синаптической передаче.
- 1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) получили Нобелевскую премию за открытие роли норадреналина в синаптической передаче.
Распространение
Электрические синапсы встречаются по всей центральной нервной системе. Они были детально изучены в неокортексе, гиппокампе, , голубом пятне, , , обонятельных луковицах, сетчатке и спинном мозге позвоночных. Электрические синапсы были обнаружены также в полосатых телах, мозжечке и супрахиазматическом ядре. Как правило, электрические синапсы опосредуют очень быстрые поведенческие ответы, как, например, выпускание чернил морским зайцем Aplysia при опасности. Электрические синапсы были обнаружены и вне нервной системы. Электрические синапсы связывают , кардиомиоциты, гладкомышечные клетки, клетки печени, глиальные и эпителиальные клетки.
Электрические синапсы наиболее характерны для низкоорганизованных животных. В ходе эволюции доля электрических синапсов уменьшалась, и в ЦНС млекопитающих (в том числе человека) на долю электрических синапсов приходится около 1 % связей между нейронами.
Функции
Морской заяц Aplysia californica выпускает чернила в качестве защитной реакции. Она опосредована электрическими синапсами
Простота устройства электрических синапсов позволяет им проводить сигнал очень быстро, однако они участвуют лишь в простых поведенческих реакциях, в отличие от более сложно устроенных химических синапсов. Поскольку для передачи сигнала через электрический синапс не нужно связывание рецептора с сигнальной молекулой-лигандом, при работе электрических синапсов не происходит задержки, которая у химических синапсов может составлять от 0,5 до 4 миллисекунд. Однако у млекопитающих различия в скоростях проведения сигнала электрическим и химическим синапсом не различаются так сильно, как у холоднокровных животных. Благодаря высокой скорости проведения сигнала электрическим синапсам несколько соседних нейронов развивают потенциал действия практически одновременно. Ответ постсинаптического нейрона имеет тот же знак, что и изменения в пресинаптическом нейроне. Так, деполяризация пресинаптической мембраны всегда вызовет деполяризацию постсинаптической мембраны, то же самое имеет место для . Как правило, ответ постсинаптического нейрона меньше, чем амплитуда исходного сигнала; это обусловлено сопротивлением пре- и постсинаптической мембран. Электрические синапсы имеют сравнительно низкую утомляемость и очень устойчивы к изменениям внешней и внутренней среды. Обычно сигнал может проходить через электрические синапсы в обоих направлениях, однако из этого правила есть исключения. Иногда в ответ на деполяризацию в мембране аксона открываются потенциалзависимые ионные каналы, которые не дают сигналу распространяться в обоих направлениях. Имеются свидетельства своего рода «пластичности» электрических синапсов, то есть электрическая связь между двумя нейронами может ослабляться или усиливаться в зависимости от активности синапса или при изменении внутриклеточной концентрации магния.