Сенсорные зоны коры
Содержание:
Ассоциативные зоны
Ассоциативные зоны находятся в коре, покрывающей большие полушария, не связаны с двигательными процессами или сенсорной деятельностью. Эти зоны головы занимают около 80% площади коры, преимущественно локализуются в долях – лобной и височной, а также затылочной и теменной. Каждый ассоциативный участок тесно взаимодействует с проекционными зонами, в том числе сенсорными и моторными, пролегающими в коре больших полушарий, образующих головной мозг.
Считается, что в этих отделах происходит объединение разрозненных фрагментов информации, в результате чего образуются сложные формы сознания. Проекционный участок опоясан ассоциативными областями, что обеспечивает взаимодействие, которое осуществляется посредством нейронов полисенсорной природы. Нервные клетки воспринимают сигналы, поступающие от разных органов и систем.

Они реагируют на информацию, передаваемую органами зрения и слуха, а также кожными рецепторами. Способность воспринимать информацию разного плана позволяет интегрировать данные, объединять их в общую систему, координировать двигательную и сенсорную деятельность. С ассоциативными зонами головного мозга и их функциями удобнее знакомиться при помощи таблицы.
| Отделы | Функции | Последствия поражения |
| Лобный | Высшая психическая деятельность – личностные характеристики, творчество, влечения | Утрата способности планировать и предвидеть, нарушение продуманного, целенаправленного поведения |
| Теменной | Формирование субъективной оценки окружающего пространства, получение представления о положении собственного тела и его перемещении | Утрата способности узнавать знакомые предметы при сохранении функциональности органов зрения |
| Височный | Речевая функция, распознавание и хранение речевой информации, воспроизводимой человеком и услышанной им | Утрата способности понимать чужую речь, невозможность распознавать слова при сохранении функциональности органов слуха |
Ассоциативные отделы, находящиеся в теменной доле, объединяют информацию, которая приходит от соматосенсорной системы. Соматосенсорная система образована рецепторами, обеспечивающими чувствительность, и центрами обработки информации, поддерживает сенсорные модальности, как температура, осязание, проприоцепция (ощущение своего тела – его положения, передвижения его частей), ноцицепция (физиологическая боль).
Ассоциативные отделы, находящиеся в височной части, отвечают за распознавание мелодий и разных сочетаний музыкальных звуков. Поле 37 позволяет запоминать слова. В височной части также содержатся центры сна, сновидений и памяти. Поле 39, расположенное на границе, разделяющей доли – височную, теменную, затылочную, содержит центр чтения, благодаря которому человек понимает письменный текст.
Визуальная зона V4[править]
Визуальная зона V4 является одной из зрительных зон в extrastriate (затылочная часть мозга) зрительной коры. Extrastriate cortex является областю затылочного cortexа (область мозжечка, охватывает левое и правое полушарие мозга) мозга млекопитающих, расположенном рядом с первичной зрительной корой, которую также называют полосатой корой из-за его полосатго вида при появлении в микроскопе. Extrastriate cortex включает в себя несколько функциональных областей, в том числе V3, V4, V5/MT, которые чувствителльны к движению, т.е. extrastriate область тела (EBA) используются в восприятии человеческих тел. В макаке она находится на передней V2 и кзади posterior нижневисочной области (PIT). Она включает в себя не менее четырех регионов (слева и справа V4d, слева и справа V4v), а некоторые группировки сообщают о том, что она содержит ростральные и хвостатые подразделения. Неизвестно функция у человека гомолога (с соразмерной, однозначной областью) зоны V4, и эта проблема является в настоящее время предметом пристального внимания.
V4-это третья область коры вентрального потока. Получив сильный прямой вход из V2 из неё идёт отправка прочных связей с ПИТ. Она также получает прямые входы от V1, особенно из Центральной площади. Кроме того, она имеет более слабые соединения и с V5 спинного prelunate (угловая извилина в теменной доле мозга) извилиной (DP).
V4 — это первая область вентрального потока для демонстрации сильного внимания вопроса модуляции
Большинство исследований указывают на то, что избирательное внимание можете изменить значимость цены исследований в зоне V4, примерно, на 20%. Основополагающей бумагой Моран и Desimone, характеризующих эти эффекты, был первый документ, о нахождения внимания на эффекты в любом месте в зрительной коре головного мозга,
Как V1, V4 зоны настроены для ориентации, определения пространственнаой частоты, и цвета. В отличие от V1, зона V4 настроена для объектов средней сложности, как на простые геометрические фигуры, хотя никто еще не разработал полныый перечень параметрического описания настройки пространства для зоны V4. Визуальная зона V4 не настроена для сложных объектов, таких как лица, таких областей в нижневисочном cortexе .
Работа свойства зоны V4 была впервые описана Semir Zeki в конце 1970-х годов, который также назвал эти области. До этого, V4 была известна её анатомическим описанием, как prelunate (теменная часть мозга) извилины. Первоначально, Зеки утверждали, что цель V4 было обрабаботать информацию о цвете. В начале 1980-х годов он доказал, что изучение зоны V4 была непосредственно связано с формой признания, как и раньше областей коры головного мозга. Это исследование поддерживает два потока гипотез, впервые представленных Ungerleider и Мишкин в 1982 году.
Недавние работы показали, что зона V4 на экспонатах долгосрочной пластичности, кодирует стимул рельефностью, выведенный на сигналы, поступающие от лобовой части глазных полей и показывает изменения в пространственных профилях его рецептивного поля внимания.
функции
Чтобы описать и понять функцию затылочной доли, следует иметь в виду, что различные области, которые составляют кору головного мозга, не имеют единой активности..
На самом деле, разные доли коры головного мозга по-разному участвуют в множественной деятельности мозга.
Несмотря на этот фактор, который определяет функционирование верхних отделов головного мозга, функция, которая лучше всего описывает деятельность затылочной доли, — это обработка визуальной информации..
На самом деле, основной функцией этой области коры головного мозга является получение стимулов, связанных с зрительным путем, которые исходят главным образом от зрительных нервов и, во вторую очередь, от других подкорковых структур..
В этом смысле затылочная доля включает в себя зрительную кору, которая является областью коры головного мозга, в которую сначала попадает информация, поступающая из сетчатки глаз и зрительных нервов..
Аналогично, зрительная кора затылочной доли делится на различные области, которые классифицируются в соответствии с уровнем обработки, которую они принимают..
Таким образом, первичная зрительная кора является частью затылочной доли, которая отвечает за обработку наиболее «сырых» визуальных данных, и является областью, ответственной за обнаружение общих закономерностей, которые можно найти в визуальной информации, собранной глазами..
Общие данные, собранные первичной зрительной корой затылочной доли, обычно не очень подробны и обычно не содержат конкретной информации о захваченном стимуле..
Впоследствии первичная зрительная кора отвечает за отправку собранной информации в другие области затылочной доли, которые отвечают за более тонкую обработку зрения..
Аналогично, другие структуры затылочной доли отвечают за отправку анализируемой информации в другие структуры мозга..
Вкратце, затылочная доля содержит области или нервные центры, которые регулируют, главным образом, следующие виды деятельности:
- Развитие мысли и эмоций.
- Интерпретация изображений.
- вид.
- Пространственная разведка.
- Различение движения и цвета.
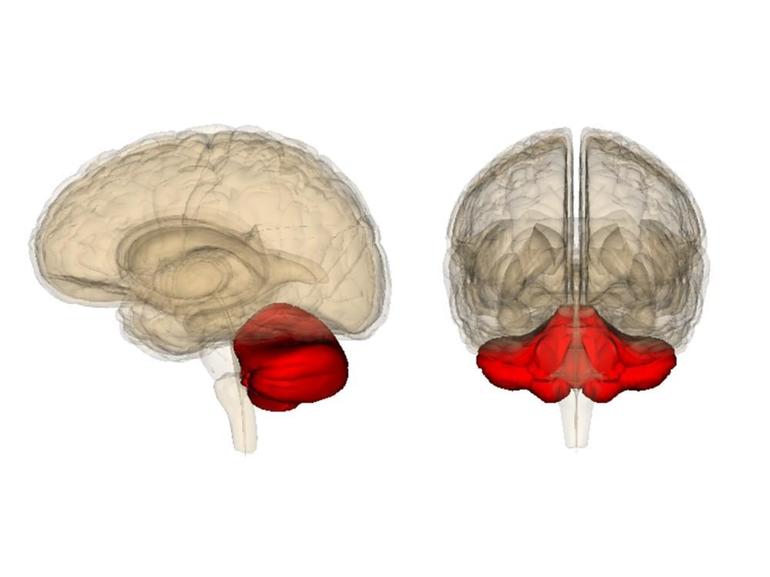
Что такое таламус
Волокна от таламических ядер (одного или нескольких) подходят к каждой доле полушарий нашего мозга. Зрительный бугор, или таламус, находится в переднем мозге, в его центральной области. Он состоит из множества ядер, при этом каждое из них передает импульс в строго определенный участок коры.
Все сигналы, поступающие к ней (кроме обонятельных), проходят сквозь релейные и интегративные ядра таламуса. Далее волокна идут от них к сенсорным зонам (в теменной доле – к вкусовой и соматосенсорной, в височной – к слуховой в затылочной – к зрительной). Поступают импульсы соответственно от вентро-базального комплекса, медиального и латерального ядер. Что касается моторных зон коры, они имеют связь с вентролатеральным и передним вентральным ядрами таламуса.
Визуальная зона V5, известная как визуальная зона MT[править]
Визуальная зона V5, также известна как визуальная зона MT (средне-височная), области extrastriate (включает в себя несколько функциональных областей, в том числе V3, V4, V5/MT, которые чувствительны к движению, или области тела (EBA), используемые в восприятии человеческих тел) зрительной коры головного мозга, которые, как полагают, играют важную роль в восприятие движения, интеграции локального движения, сигналов с мировыми идеями и роль в руководстве некоторыми движениями глаз.
Соединенияправить
Рис.ТЗБ. Теменно-затылочная борозда
MT подключен к огромному массиву корковых и подкорковых областей мозга. Его входы включают в себя визуальные области коры головного мозга V1, V2, и спинную область V3 (dorsomedial курения — область известноя как DM или V6, подразделение зрительной коры приматов. DM находится в дорсальной части extrastriate cortex, рядом с глубокой канавкой, проходящей через центр мозга (межполушарной щели), и, как правило, также включает в медиальной части коры головного мозга, такие области как Parieto-затылочной sulcus — теменно-затылочную борозду (см. рис.ТЗБ) (sulcus parieto-occipitalis)),, в koniocellular регионы LGN, и уступает подушке зрительного бугра. Шаблон прогнозов на МТ несколько меняется между представлениями центрального и периферического поля зрения, причем последние получают ресурсы от участков, расположенных в средней линии и коре retrosplenial области
Стандартное представление, что V1 обеспечивает «наиболее важные» входы на Афон. Тем не менее, несколько исследований показали, что нейроны в MT реагирует на визуальную информацию, часто не в том направлении, — селективным образом, но даже после зоны V1 были уничтожены или обезврежены. Кроме того, научно-исследовательская работа Semir Zeki и коллаборационистов предположили, что определенные виды визуальной информации может достигать при помощи MT, прежде чем она достигает даже в зоне V1.
MT отправляет его основные выходы в регионы, расположенные в коре головного мозга, непосредственно окружающеие, в том числе в районы FST, Минспорта и V4t (средней височной Полумесяца). Другие прогнозы MT — ставят цель исследования движения глаз в областях, связанных с лобной и теменной долями (лобной области глаз и боковых внутрстеночных областей).
Функцияправить
Первые исследования электрофизиологических свойств нейронов в MT показали, что большая часть клеток были tuned (Нейрональный тюнинг относится к собственности клеток головного мозга, с помощью которых они избирательно представляют определенный тип сенсорного, моторного, или роль в познавательной информации) скорости и направление движения визуальных раздражителей., Эти результаты позволяют предположить, что МТ сыграл значительную роль в обработке зрительных движения.
Отрицательные исследования также поддержали роль МП в восприятии движения и движения глаз. Нейропсихологические исследования больного, который не мог видеть движения, видя мир в серии статических «кадров» высказало мнение о том, что МТ в приматах — это аналогично гомологичным областям V5 у человека.,
Однако, поскольку нейроны в зоне V1 также выверенное направление и в скорости движения, эти предварительные результаты оставляют открытым вопрос, какой именно МТ могли бы сделать, что бы не быть зоной V1. Большая работа была проведена в этом регионе, как представляется должны быть, чтобы интегрировать местные visual движения сигналов в мировом движении сложных объектов. Например, необходимо зону V5 привести к дефициту в восприятии движения и обработки сложных раздражителей. МТ содержит много нейронов с избирательными движениями со сложными визуальными характеристиками (концов линии, углов). Microstimulation нейрона, расположенном в зоне V5 влияет на восприятие движения. Например, если кто-то находит нейрон с предпочтением для восходящего движения, и тогда мы с помощью электродов, чтобы это стимулировать, вызывает у обезьяна склонность сообщать движения вверх.
Есть еще много споров по точной формы расчетов, проведенных в области МТ и некоторые исследования показывают, что есть движение, по сути, уже доступные на более низких уровнях зрительной системы, такие как в зоне V1.,
Центральные поля
Ученые за долгие годы изучения накопили значительный опыт клинических исследований. В результате наблюдений было установлено, в частности, что повреждения тех или иных полей в составе корковых представителей анализаторов сказываются на общей клинической картине далеко не равнозначно. Среди остальных полей в этом отношении выделяется одно, которое в ядерной зоне занимает центральное положение. Оно называется первичным, или центральным. Им является поле под номером 17 в зрительной зоне, в слуховой – под номером 41, а в кинестетической – 3. Их повреждение ведет к очень серьезным последствиям. Теряется способность воспринимать или осуществлять самые тонкие дифференцировки раздражителей соответствующих анализаторов.
Структура коркового слоя и функции отделов
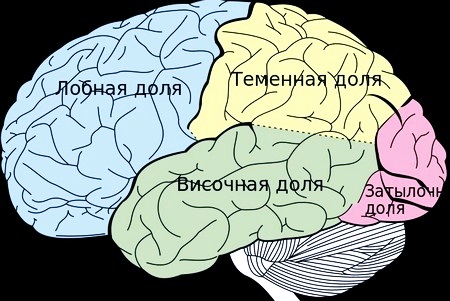
Продольная борозда разделяет мозг на большие полушария, которые состоят из 6 функциональных зон коры:
- Лобная.
- Теменная.
- Височная.
- Затылочная.
- Островок. Находится в углублении в Сильвиевой борозде.
- Лимбическая. Располагается с краю каждого полушария относительно срединной плоскости.
Несмотря на относительное разделение функций отдельных зон коры головного мозга, каждый физиологический процесс, протекающий в организме, требует их тесного взаимодействия и предполагает функциональную интеграцию. К примеру, зрительный центр располагается в затылочной области, однако в комплексном восприятии и обработке зрительного стимула кроме затылочной доли участвуют лобные и височные отделы.
К примеру, левое доминирует в формировании речи, правое руководит в процессе обеспечения ориентации в пространстве. Первичные зоны коркового слоя головного мозга бывают моторными и сенсорными, другие отделы называют ассоциативными, которые еще известны как унимодальные и гетеромодальные. Унимодальные отделы находятся рядом с соответствующей сенсорной областью.
Они более тонко и глубоко обрабатывают информацию, поступающую в сенсорную область. Гетеромодальные отделы получают конвергентные (обладающие схожими признаками) данные из множества сенсорных и двигательных отделов. Благодаря работе гетеромодальных отделов происходит сравнение вновь поступающей информации с данными, хранящимися в инстинктивной и приобретенной памяти.
Корковый слой мозга разделен на зоны, которые в зависимости от расположения отвечают за выполнение разных задач, что позволяет выявлять участок поражения нервной ткани на основании симптомов. К примеру, зона Брока, известная как двигательный речевой центр, находится в височной части коркового слоя. Повреждение этого участка мозга провоцирует развитие моторной афазии. Пациент понимает речь, но не способен произносить слова, разговаривать.
Осуществляемые корой функции часто обуславливаются устройством структуры. Строение коры головного мозга отличается своими особенностями, которые выражаются в разном количестве слоев, размерах, топографии и строении формирующих кору нервных клеток. Ученые различают несколько разных видов слоев, которые, взаимодействуя друг с другом, способствуют функционированию системы полностью:
- молекулярный слой: он создает большое количество хаотичным образом сплетенных дендритных образований с небольшим содержанием клеток, по форме похожих на веретено, которые отвечают за ассоциативное функционирование;
- внешний слой: выражен большим числом нейронов, которые имеют разнообразную форму и высокое содержание. За ними расположены внешние пределы структур, по форме напоминающие пирамиду;
- внешний слой пирамидального вида: содержит в себе нейроны незначительных и существенных габаритов во время более глубокого нахождения больших. По форме эти клетки напоминают конус, от верхней точки отходит дендрит, который имеет максимальные габариты, посредством разделения на мелкие образования связываются нейроны, содержащие серое вещество. По мере приближения к коре полушарий, ветвления отличаются небольшой толщиной и формируют структуру, напоминающую по форме веер;
- внутренний слой зернистого вида: содержит в себе нервные клетки, которые имеют маленький размер, располагаются на определенном расстоянии, между ними идут сгруппировавшиеся структуры волокнистого вида;
- внутренний слой пирамидального вида: включает в себя нейроны, которые обладают средними и большими габаритами. Верхние окончания дендритов могут доходить до молекулярного слоя;
- покров, который содержит в себе нейронные клетки, обладающие формой веретена. Свойственно для них то, что их часть, которая находится в самой низкой точке, может достигнуть уровня белого вещества.
Строение коры большого мозга довольно сложное и обладает своими особенностями, выражающимися в разном количестве покровов, габаритов, топографии и структуре клеток, которые образовывают слои.
Первичная зрительная кора (V1)
Рис.4. Мозг человека.Красным цветом обозначена первичная зрительная кора (зрительная зона V1)
Первичная зрительная кора — наиболее изученная зрительная зона мозга. Исследования показали, что у млекопитающих она занимает задний полюс затылочной доли каждого полушария (эти доли ответственны за обработку зрительных стимулов). Это наиболее просто устроенная и филогенетически более «древняя» из кортикальных зон, связанных со зрением. Она приспособлена для обработки информации о статических и движущихся объектах, в особенности, для распознавания простых образов.
Составная часть функциональной архитектуры коры больших полушарий головного мозга — первичная зрительная кора — практически полностью соответствует анатомически определяемой стриарной коре. Название последней восходит к латинскому «полоса, полоска» (лат. stria) и во многом обусловлено тем, что здесь отчётливо видна невооружённым глазом полоска Дженнари (наружная полоска Байярже), образованная конечными отделами покрытых миелиновой оболочкой аксонов, отходящих от нейронов латерального коленчатого тела и заканчивающихся в IV слое серого вещества.
Первичная зрительная кора подразделяется на шесть функционально различающихся горизонтальных цитоархитектонических слоёв, обозначаемых римскими цифрами от I до VI.
Слой IV (внутренний зернистый слой), к которому подходит наибольшее количество афферентных волокон, идущих от латеральных коленчатых тел (ЛКТ), в свою очередь, подразделяется на четыре подслоя, обозначаемых IVA, IVB, IVCα и IVCβ. Нервные клетки подслоя IVCα, в основном, получают сигналы, идущие от нейронов магноцеллюлярных («крупноклеточных», вентральных) слоёв ЛКТ («магноцеллюлярный зрительный путь»), подслоя IVCβ — от нейронов парвоцеллюлярных («мелкоклеточных», дорсальных) слоёв ЛКТ («парвоцеллюлярный зрительный путь»).
Полагают, что среднее число нейронов первичной зрительной коры взрослого человека составляет около 140 миллионов в каждом полушарии.
Примечания
- ↑
- ↑ Часть III. Общая и специальная сенсорная физиология // Физиология человека: в 3-х томах = Human Physiology. Ed. by R.F. Schmidt, G. Thews. 2nd, completely revised edition (translated from German by M.A. Biederman-Thorson) / под ред. Р. Шмидта и Г. Тевса. — изд-е 2-е, перераб. и дополн.. — М.: Мир, 1996. — Т. 1. Пер. с англ.. — С. 178—321. — 323 с. — 10 000 экз. — ISBN 5-03-002545-6.
- Rita L. Atkinson, Richard C. Atkinson, Edward E. Smith, Daryl J. Bem, Susan Nolen-Hoeksema. «Hilgard’s Introduction to Psychology. History, Theory, Research, and Applications», 13th ed., 2000
- ↑
- Ungerleider L. G., Mishkin M. Two Cortical Visual Systems // Analysis of Visual Behavior (неопр.) / Ingle D. J., Goodale MA and Mansfield RJW. — Boston: MIT Press, 1982. — С. 549—586.
- Королькова О. А., Печенкова Е. В., Синицын В. Е. ФМРТ-исследование функциональных связей головного мозга в задаче категоризации эмоциональных и нейтральных лиц/Когнитивная наука в Москве: новые исследования. Материалы конференции 15 июня 2017 г. Под ред. Е.В. Печенковой, М.В. Фаликман. – М.: ООО «Буки Веди», ИППиП. 2017 г. – 596 стр. Электронная версия. С.174-179. ISBN 978-5-4465-1509-7
- Pitcher D., Walsh V., Duchaine B. The role of the occipital face area in the cortical face perception network // Experimental Brain Research. 2011. Vol. 209. No. 4. P. 481 – 493. doi:10.1007/s00221-011-2579-1
- Kanwisher N., McDermott J., Chun M. M. The fusiform face area: a module in human extra- striate cortex specialized for face perception // Journal of Neuroscience. 1997. Vol. 17. No. 11. P. 4302 – 4311
- HaxbyJ.V., HoffmanE.A., GobbiniM.I. The distributed human neural system for face perception // Trends in Cognitive Sciences. 2000. Vol. 4. No. 6. P. 223 – 233. doi:10.1016/ s1364-6613(00)01482-0
- Ishai A. Let’s face it: It’s a cortical network // NeuroImage. 2008. Vol. 40. No. 2. P. 415 – 419. doi:10.1016/j.neuroimage.2007.10.040
- Синельников Р.Д., Синельников Я.Р. Атлас анатомии человека: в 4-х томах. — изд-е 2-е, стереотипное. — М.: Медицина, 1996. — Т. 3. — 232 с. — (Учеб. лит. для студ. мед. вузов). — ISBN 5-225-02722-9.
- Фениш Х. Карманный атлас анатомии человека на основе Международной номенклатуры = Feneis H. Pocket Atlas of Human Anatomy Based on International Nomenclature. — NY, 1994 / редактор перевода канд. мед. наук, проф. С.Д. Денисов. — 3-е изд., стереотип.. — Минск: Выш.шк., 2000. — С. 376—377. — 464 с. — 10 000 экз. — ISBN 985-06-0592-8.
- Хьюбел Д., Т. Визель. Центральные механизмы зрения // Мозг (перевод с английского специального выпуска журнала Scientific American, 1979) / Редактор перевода П. В. Симонов. — М.: Мир, 1982. — 279 с. — 15 000 экз.
- ↑ Быков В. Л. Кора полушарий большого мозга // Частная гистология человека. — СПб.: СОТИС, 2001. — С. 260—271. — 304 с. — 3 000 экз. — ISBN 5-85503-116-0.
- ↑ Хьюбел Д. Глаз, мозг, зрение = Eye, Brain and Vision / Под редакцией чл.-корр. АН СССР А. Л. Бызова. — М.: Мир, 1990. — 239 с. — 48 000 экз. — ISBN 5-03-001254-0.
- ↑
- ↑
Визуальная зона V2[править]
Рис.6.В feed forward сети информация всегда движется в одном направлении; она никогда не пойдет в обратном направлении.
Визуальная зона V2, которая также называется (prestriate cortex или область, которая включает в себя визуальные зоны: V2,V3,V4,MT (иногда называемые (V5), и визуальную зону DP.), является вторым важнейшим направлением в зрительной коре, и первым регионом в системе зрительных ассоциаций «курения». Она получает мощную функцию (feedforward или движение информации только в одном направлении, вперед, от входных узлов, через скрытые узлы (если таковые имеются) и от выходных узлов. Нет циклов и петель в сети) — нейронную сеть прямого распространения — по типу искусственных нейронных сетей, где связь между подразделениями не сформировать направленем цикла. Соединения от зоны V1 непосредственно и через подушку зрительного бугра и отправляет информацию посредством прочных связей с V3, V4 и V5. Визуальная зона V2 также имеет сильную обратную связь с подключениями к зоне V1.
Анатомически, V2 разделена на четыре квадранта, дорсальный’ и вентральный в левой и правой частях полушарий. Вместе эти четыре региона, обеспечивая полную карту визуального мира. Функционально, V2 имеет много общих свойств с V1. Клетки настроены к простым свойствам: таких как выбор ориентации, пространственная частота, и цвет. Ответы на многие V2 нейроны и модуляция более сложных свойств, таких как ориентация иллюзорных контуров, бинокулярное неравенство, и будет ли стимулироваться часть изображения или земли (Цю и фон дер Heydt, 2005). Недавние исследования показали, что V2 клетки показывают небольшое количество внимания в модуляции (более V1, меньше, чем V4), умеренно настроенные сложные структуры, и может быть обусловлены несколькими ориентациями в различных субрегионах в пределах одного рецептивное поля.
Цю и фон дер Heydt, 2005 утверждал, что весь вентральной visual-гиппокампа поток важен для зрительной памяти. Эта теория, в отличие от доминирующей, предсказывает, что объект-признание памяти (ORM) изменения, могущие произойти в результате манипуляций в V2, в районах, которые сильно связаны между собой в пределах вентрального потока зрительной коры. В обезьянем мозгу, эта область получает сильные прямые соединения из первичной зрительной коры (V1) и посылает сильные прогнозы на другие вторичные зрительные коры (V3, V4 и V5). Большинство нейронов этой области настроены на простых визуальных характеристик, таких как выбор ориентации, пространственная частота, размер, цвет и форма.,, V2 клетки также реагировать на различные комплексы характеристик формы, такие как ориентация иллюзорных контуров и будет ли стимул части рисунка или земли. Анатомические исследования предполагают layer 3 V2 в области визуальной обработки информации. В отличие от layer 3 слоя (оптический слой), слой 6 зрительной коры состоит из множества типов нейронов, и их реакции на зрительные стимулы-более сложные.
В недавнем исследовании, в Слое 6 клеток V2 коры были найдены характеристики, которые играют очень важную роль в хранении, распознавании объектов памяти, а также краткосрочные превращения этих объекта воспоминания в долгосрочные воспоминания.
Третичные зоны
Функциональные отношения, которые прослеживаются между анализаторами, еще более сложны, чем было описано выше. Морфологически дальнейшее их усложнение выражается в том, что в процессе роста по поверхности полушария ядерных полей анализаторов эти зоны взаимно перекрываются. У корковых концов анализаторов образуются «зоны перекрытия», то есть третичные зоны. Данные формации относятся к самым сложным типам объединения деятельности кожно-кинестетического, слухового и зрительного анализаторов. Третичные зоны расположены уже за границами собственных ядерных полей. Поэтому их раздражение и повреждение не приводит к выраженным явлениям выпадения. Также и в отношении специфических функций анализатора не наблюдаются значительные эффекты.
Третичные зоны – это особые области коры. Их можно назвать собранием «рассеянных» элементов различных анализаторов. То есть это элементы, которые сами по себе уже не способны производить какие бы то ни было сложные синтезы или анализы раздражителей. Территория, которую они занимают, достаточно обширна. Она распадается на целый ряд областей. Вкратце опишем их.
Верхняя теменная область важна для интеграции движений всего тела со зрительными анализаторами, а также для формирования схемы тела. Что касается нижней теменной, то она относится к объединению отвлеченных и обобщенных форм сигнализации, связанных со сложно и тонко дифференцированными речевыми и предметными действиями, выполнение которых контролируется зрением.
Область височно-теменно-затылочная также очень важна. Она отвечает за сложные типы интеграции зрительного и слухового анализаторов с письменной и устной речью.
Отметим, что третичные зоны имеют самые сложные цепи связи по сравнению с первичными и вторичными. Двусторонние связи наблюдаются у них с комплексом ядер таламуса, связанными, в свою очередь, с реле-ядрами посредством длинной цепи внутренних связей, имеющихся непосредственно в таламусе.
На основании вышеизложенного ясно, что у человека зоны первичные, вторичные и третичные представляют собой участки коры, являющиеся высоко специализированными. Особенно нужно подчеркнуть, что 3 группы корковых зон, описанные выше, в нормально работающем мозге вместе с системами связей и переключений между собой, а также с подкорковыми образованиями функционируют как одно сложно дифференцированное целое.